{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
EMD-2451
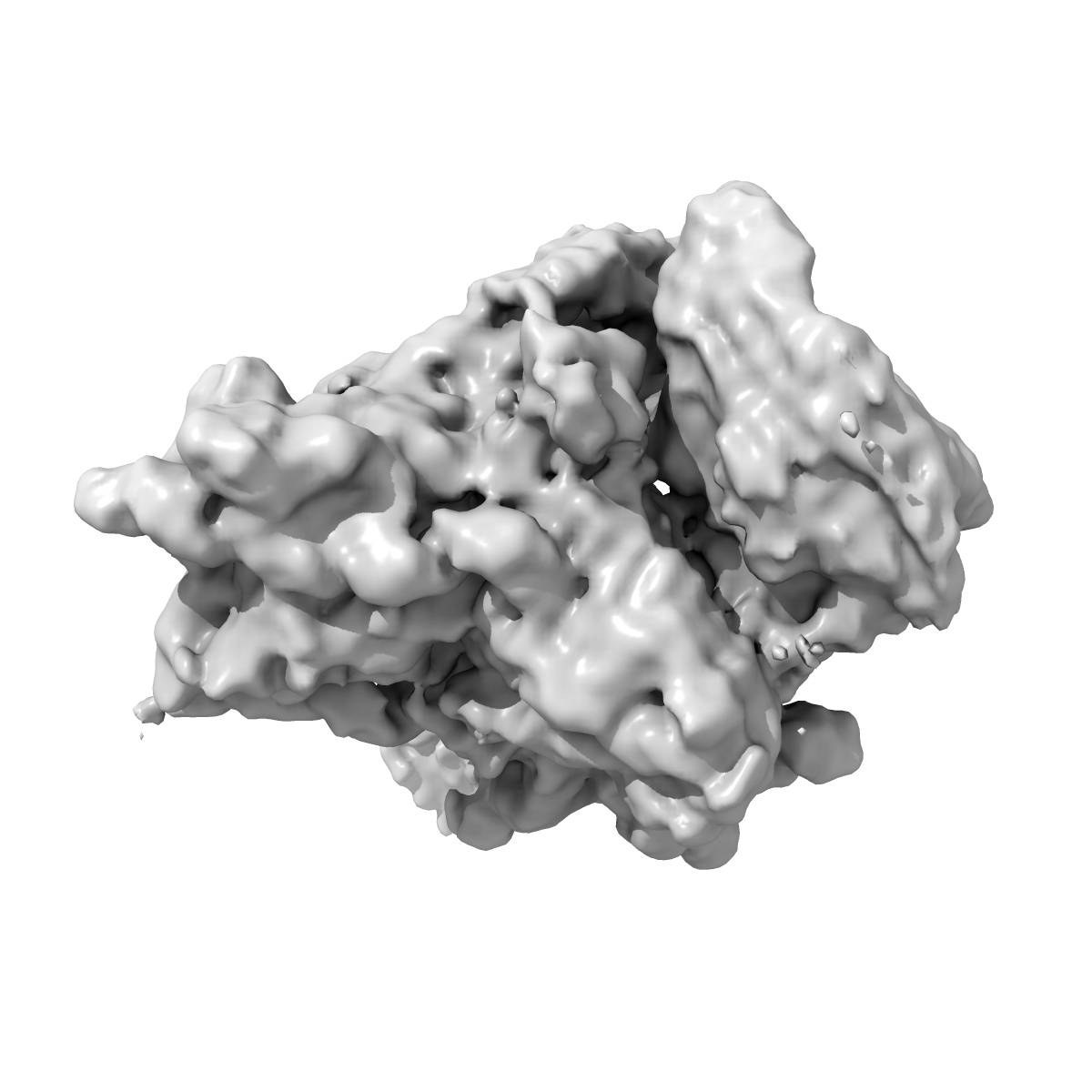
Cryo-EM structure of the CSFV IRES in complex with eIF3, small ribosomal 40S subunit and DHX29
EMD-2451
Single-particle9.3 Å
 Deposition: 07/09/2013
Deposition: 07/09/2013Map released: 13/11/2013
Last modified: 17/02/2016
Sample Organism:
Oryctolagus cuniculus,
Classical swine fever virus,
Homo sapiens
Sample: Reconstruction of a mutant Classical Swine Fever Virus IRES bound to eukaryotic initiation factor 3, the Rabbit 40S subunit and DHX29.
Deposition Authors: Hashem Y, des-Georges A, Dhote V, Langlois R, Liao HY, Grassucci RA, Pestova TV, Hellen CUT, Frank J
Sample: Reconstruction of a mutant Classical Swine Fever Virus IRES bound to eukaryotic initiation factor 3, the Rabbit 40S subunit and DHX29.
Deposition Authors: Hashem Y, des-Georges A, Dhote V, Langlois R, Liao HY, Grassucci RA, Pestova TV, Hellen CUT, Frank J
Hepatitis-C-virus-like internal ribosome entry sites displace eIF3 to gain access to the 40S subunit.
Hashem Y,
des Georges A  ,
Dhote V,
Langlois R,
Liao HY,
Grassucci RA,
Pestova TV,
Hellen CU,
Frank J
,
Dhote V,
Langlois R,
Liao HY,
Grassucci RA,
Pestova TV,
Hellen CU,
Frank J
(2013) Nature , 503 , 539 - 543
 ,
Dhote V,
Langlois R,
Liao HY,
Grassucci RA,
Pestova TV,
Hellen CU,
Frank J
,
Dhote V,
Langlois R,
Liao HY,
Grassucci RA,
Pestova TV,
Hellen CU,
Frank J
(2013) Nature , 503 , 539 - 543
Abstract:
Hepatitis C virus (HCV) and classical swine fever virus (CSFV) messenger RNAs contain related (HCV-like) internal ribosome entry sites (IRESs) that promote 5'-end independent initiation of translation, requiring only a subset of the eukaryotic initiation factors (eIFs) needed for canonical initiation on cellular mRNAs. Initiation on HCV-like IRESs relies on their specific interaction with the 40S subunit, which places the initiation codon into the P site, where it directly base-pairs with eIF2-bound initiator methionyl transfer RNA to form a 48S initiation complex. However, all HCV-like IRESs also specifically interact with eIF3 (refs 2, 5-7, 9-12), but the role of this interaction in IRES-mediated initiation has remained unknown. During canonical initiation, eIF3 binds to the 40S subunit as a component of the 43S pre-initiation complex, and comparison of the ribosomal positions of eIF3 and the HCV IRES revealed that they overlap, so that their rearrangement would be required for formation of ribosomal complexes containing both components. Here we present a cryo-electron microscopy reconstruction of a 40S ribosomal complex containing eIF3 and the CSFV IRES. Remarkably, although the position and interactions of the CSFV IRES with the 40S subunit in this complex are similar to those of the HCV IRES in the 40S-IRES binary complex, eIF3 is completely displaced from its ribosomal position in the 43S complex, and instead interacts through its ribosome-binding surface exclusively with the apical region of domain III of the IRES. Our results suggest a role for the specific interaction of HCV-like IRESs with eIF3 in preventing ribosomal association of eIF3, which could serve two purposes: relieving the competition between the IRES and eIF3 for a common binding site on the 40S subunit, and reducing formation of 43S complexes, thereby favouring translation of viral mRNAs.
Hepatitis C virus (HCV) and classical swine fever virus (CSFV) messenger RNAs contain related (HCV-like) internal ribosome entry sites (IRESs) that promote 5'-end independent initiation of translation, requiring only a subset of the eukaryotic initiation factors (eIFs) needed for canonical initiation on cellular mRNAs. Initiation on HCV-like IRESs relies on their specific interaction with the 40S subunit, which places the initiation codon into the P site, where it directly base-pairs with eIF2-bound initiator methionyl transfer RNA to form a 48S initiation complex. However, all HCV-like IRESs also specifically interact with eIF3 (refs 2, 5-7, 9-12), but the role of this interaction in IRES-mediated initiation has remained unknown. During canonical initiation, eIF3 binds to the 40S subunit as a component of the 43S pre-initiation complex, and comparison of the ribosomal positions of eIF3 and the HCV IRES revealed that they overlap, so that their rearrangement would be required for formation of ribosomal complexes containing both components. Here we present a cryo-electron microscopy reconstruction of a 40S ribosomal complex containing eIF3 and the CSFV IRES. Remarkably, although the position and interactions of the CSFV IRES with the 40S subunit in this complex are similar to those of the HCV IRES in the 40S-IRES binary complex, eIF3 is completely displaced from its ribosomal position in the 43S complex, and instead interacts through its ribosome-binding surface exclusively with the apical region of domain III of the IRES. Our results suggest a role for the specific interaction of HCV-like IRESs with eIF3 in preventing ribosomal association of eIF3, which could serve two purposes: relieving the competition between the IRES and eIF3 for a common binding site on the 40S subunit, and reducing formation of 43S complexes, thereby favouring translation of viral mRNAs.