{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
EMD-30475
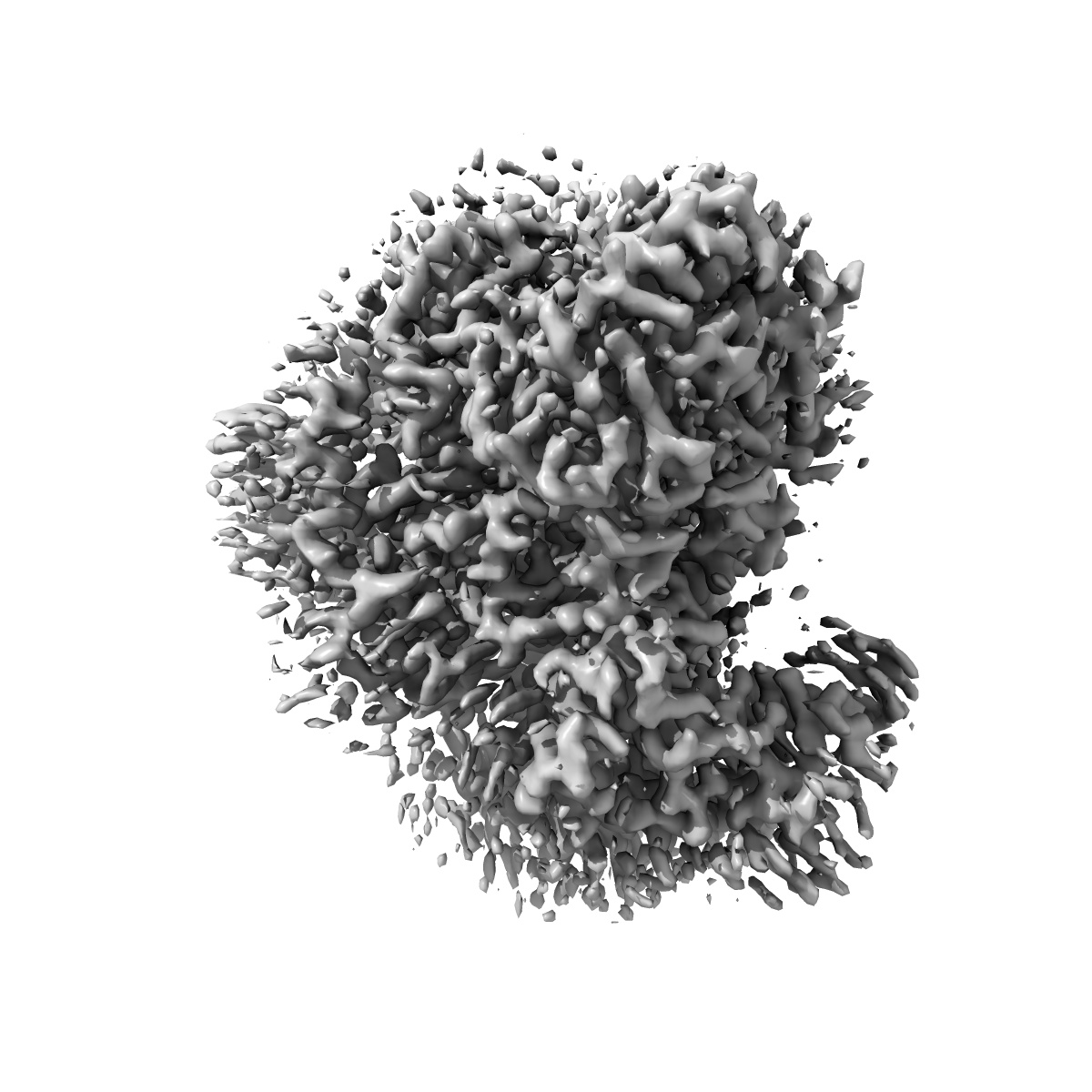
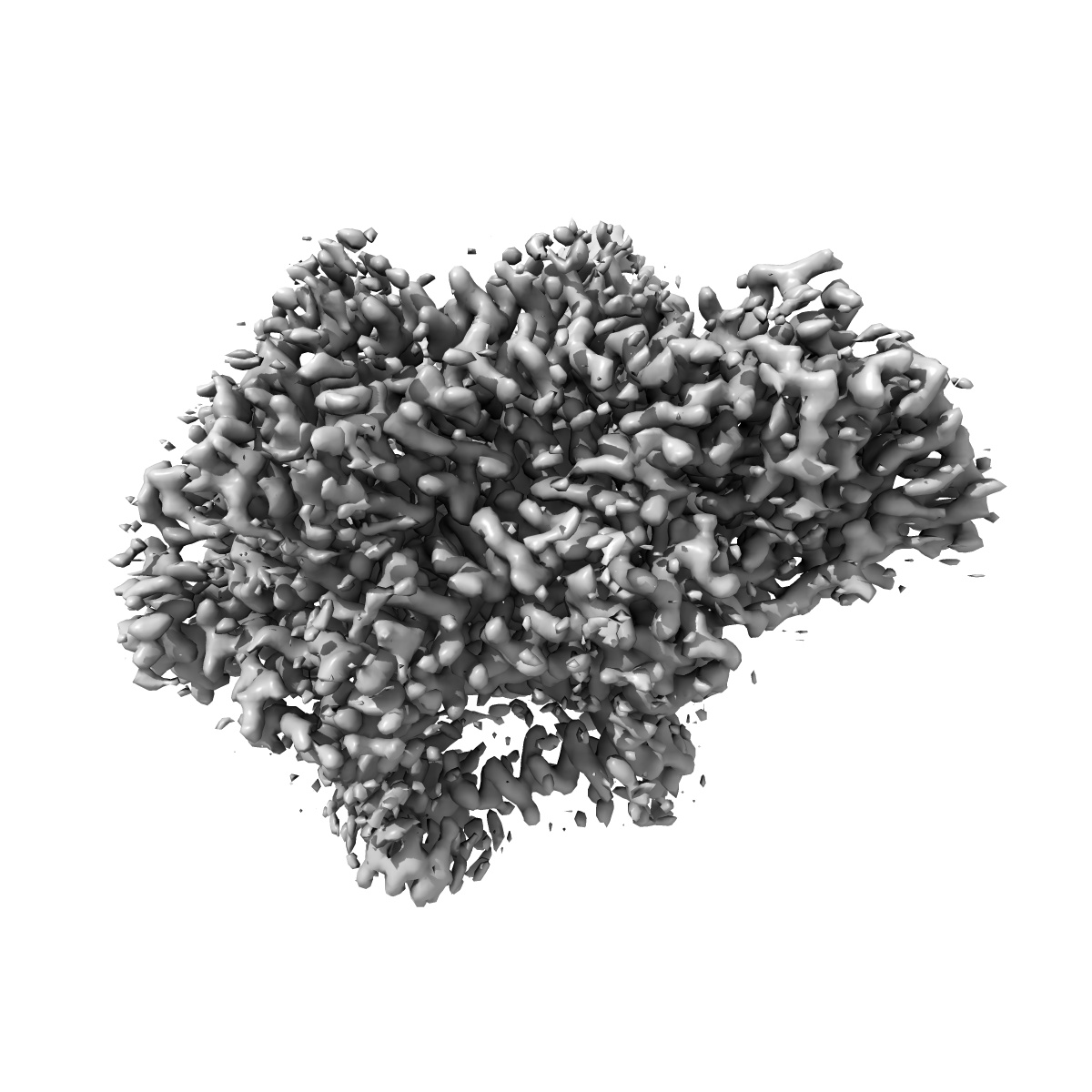
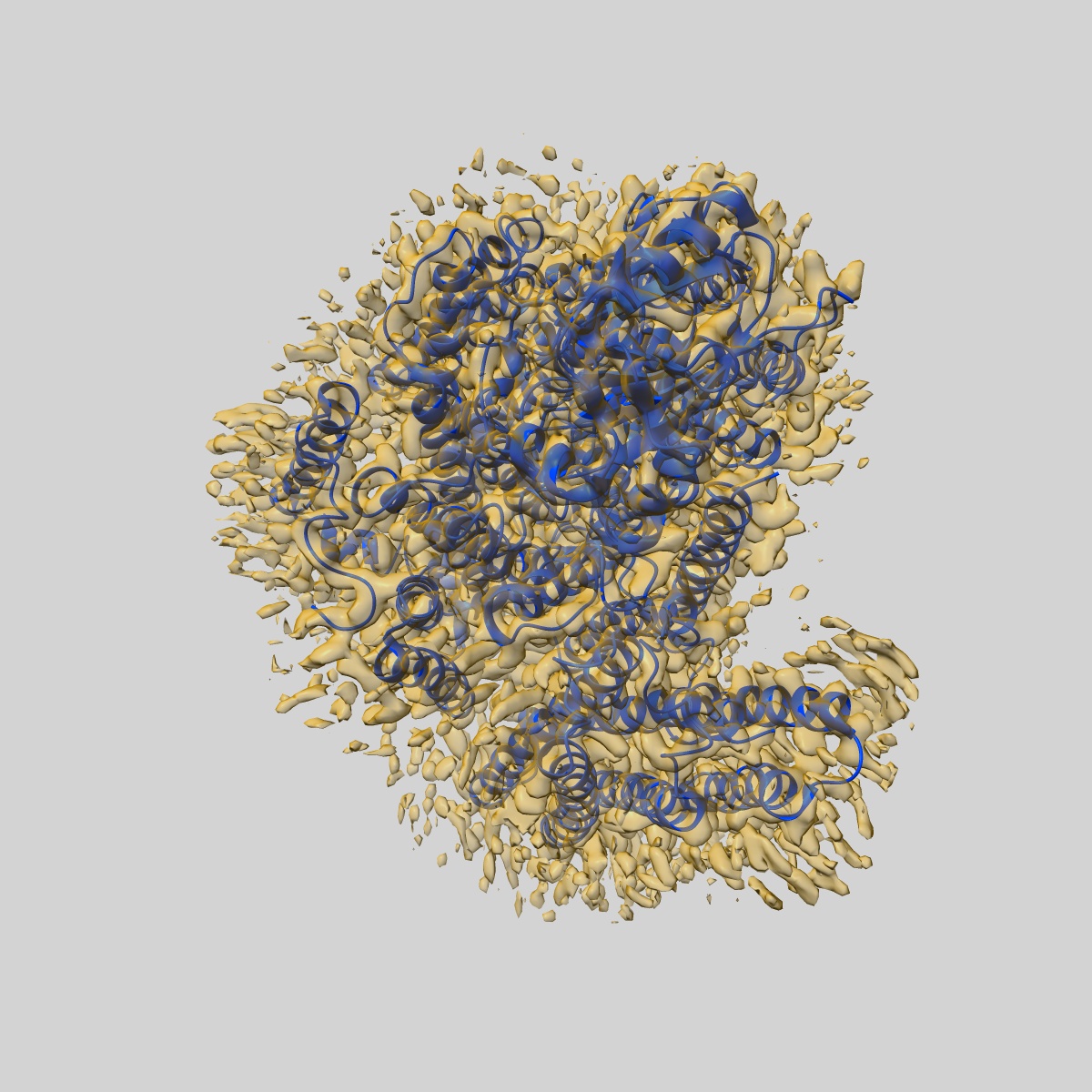
Ubiquinol Binding Site of Cytochrome bo3 from Escherichia coli
EMD-30475
Single-particle2.63 Å
 Deposition: 25/08/2020
Deposition: 25/08/2020Map released: 25/08/2021
Last modified: 29/05/2024
Sample Organism:
Escherichia coli
Sample: Cytochrome bo(3) ubiquinol oxidase
Fitted models: 7cuw (Avg. Q-score: 0.63)
Deposition Authors: Li J, Han L
Sample: Cytochrome bo(3) ubiquinol oxidase
Fitted models: 7cuw (Avg. Q-score: 0.63)
Deposition Authors: Li J, Han L

Cryo-EM structures of Escherichia coli cytochrome bo3 reveal bound phospholipids and ubiquinone-8 in a dynamic substrate binding site.
Li J,
Han L ,
Vallese F,
Ding Z ,
Choi SK ,
Hong S,
Luo Y,
Liu B,
Chan CK,
Tajkhorshid E ,
Zhu J ,
Clarke O ,
Zhang K,
Gennis R
(2021) PNAS , 118
,
Vallese F,
Ding Z ,
Choi SK ,
Hong S,
Luo Y,
Liu B,
Chan CK,
Tajkhorshid E ,
Zhu J ,
Clarke O ,
Zhang K,
Gennis R
(2021) PNAS , 118
Abstract:
Two independent structures of the proton-pumping, respiratory cytochrome bo3 ubiquinol oxidase (cyt bo3 ) have been determined by cryogenic electron microscopy (cryo-EM) in styrene-maleic acid (SMA) copolymer nanodiscs and in membrane scaffold protein (MSP) nanodiscs to 2.55- and 2.19-Å resolution, respectively. The structures include the metal redox centers (heme b, heme o3 , and CuB), the redox-active cross-linked histidine-tyrosine cofactor, and the internal water molecules in the proton-conducting D channel. Each structure also contains one equivalent of ubiquinone-8 (UQ8) in the substrate binding site as well as several phospholipid molecules. The isoprene side chain of UQ8 is clamped within a hydrophobic groove in subunit I by transmembrane helix TM0, which is only present in quinol oxidases and not in the closely related cytochrome c oxidases. Both structures show carbonyl O1 of the UQ8 headgroup hydrogen bonded to D75I and R71I In both structures, residue H98I occupies two conformations. In conformation 1, H98I forms a hydrogen bond with carbonyl O4 of the UQ8 headgroup, but in conformation 2, the imidazole side chain of H98I has flipped to form a hydrogen bond with E14I at the N-terminal end of TM0. We propose that H98I dynamics facilitate proton transfer from ubiquinol to the periplasmic aqueous phase during oxidation of the substrate. Computational studies show that TM0 creates a channel, allowing access of water to the ubiquinol headgroup and to H98I.
Two independent structures of the proton-pumping, respiratory cytochrome bo3 ubiquinol oxidase (cyt bo3 ) have been determined by cryogenic electron microscopy (cryo-EM) in styrene-maleic acid (SMA) copolymer nanodiscs and in membrane scaffold protein (MSP) nanodiscs to 2.55- and 2.19-Å resolution, respectively. The structures include the metal redox centers (heme b, heme o3 , and CuB), the redox-active cross-linked histidine-tyrosine cofactor, and the internal water molecules in the proton-conducting D channel. Each structure also contains one equivalent of ubiquinone-8 (UQ8) in the substrate binding site as well as several phospholipid molecules. The isoprene side chain of UQ8 is clamped within a hydrophobic groove in subunit I by transmembrane helix TM0, which is only present in quinol oxidases and not in the closely related cytochrome c oxidases. Both structures show carbonyl O1 of the UQ8 headgroup hydrogen bonded to D75I and R71I In both structures, residue H98I occupies two conformations. In conformation 1, H98I forms a hydrogen bond with carbonyl O4 of the UQ8 headgroup, but in conformation 2, the imidazole side chain of H98I has flipped to form a hydrogen bond with E14I at the N-terminal end of TM0. We propose that H98I dynamics facilitate proton transfer from ubiquinol to the periplasmic aqueous phase during oxidation of the substrate. Computational studies show that TM0 creates a channel, allowing access of water to the ubiquinol headgroup and to H98I.