EMD-32372
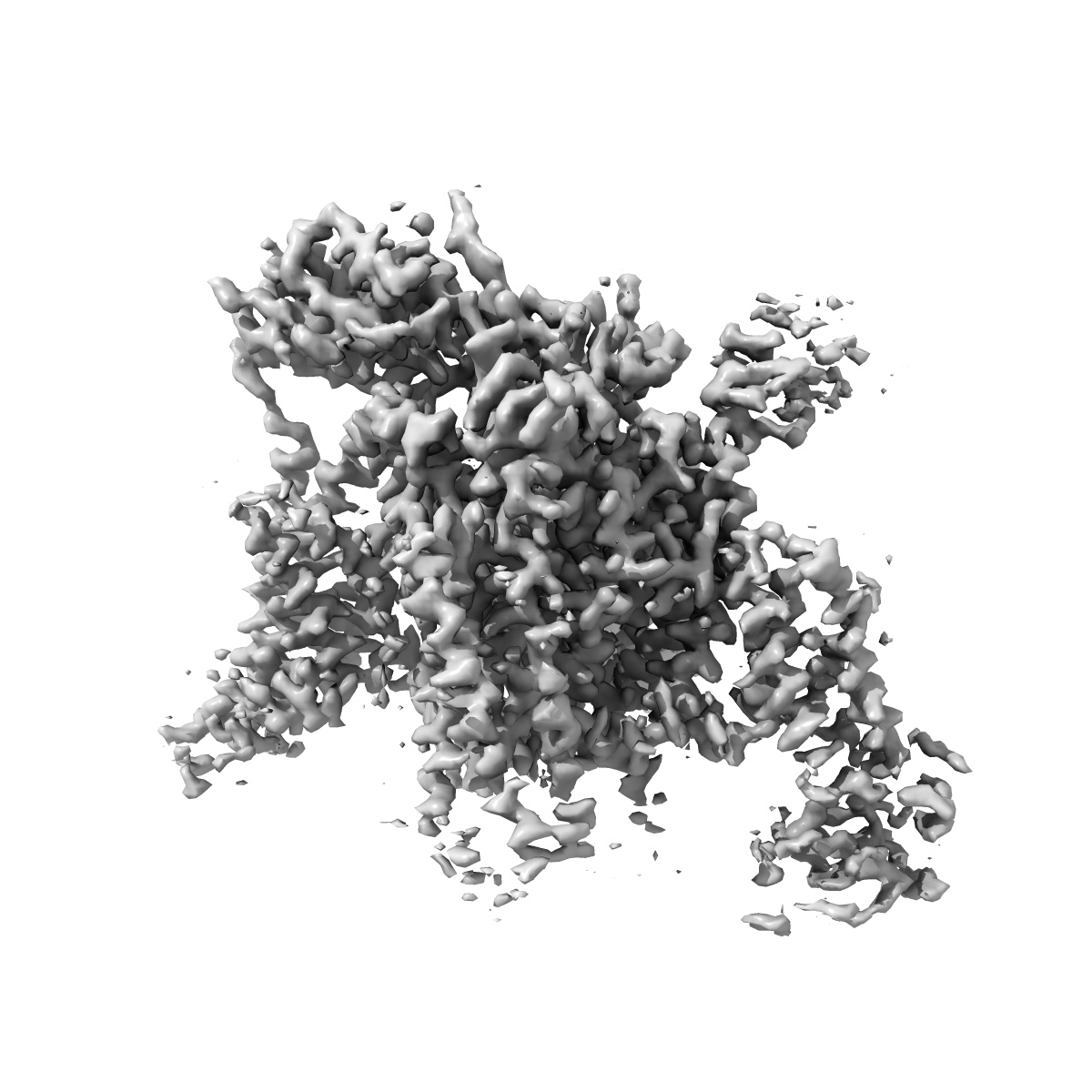
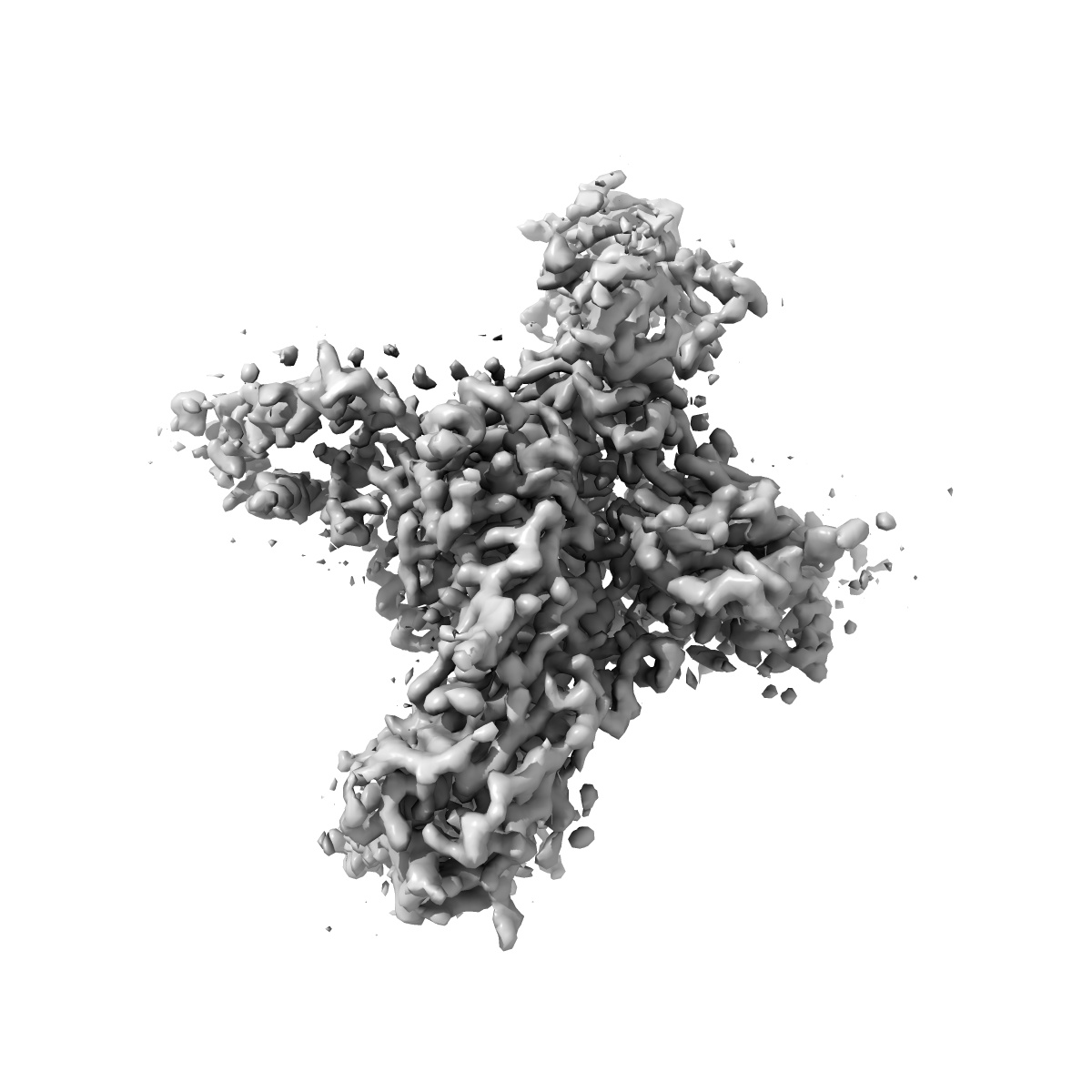
Cryo-EM structure of human Nav1.7(E406K) in complex with auxiliary beta subunits, huwentoxin-IV and saxitoxin (S6IV alpha helix conformer)
EMD-32372
Single-particle3.0 Å
 Deposition: 10/12/2021
Deposition: 10/12/2021Map released: 25/05/2022
Last modified: 30/10/2024
Sample Organism:
Homo sapiens
Sample: human Nav1.7 in complex with beta1, beta2 and Huwentoxin
Fitted models: 7w9t (Avg. Q-score: 0.431)
Deposition Authors: Yan N, Huang G, Liu D, Wei P
Sample: human Nav1.7 in complex with beta1, beta2 and Huwentoxin
Fitted models: 7w9t (Avg. Q-score: 0.431)
Deposition Authors: Yan N, Huang G, Liu D, Wei P
High-resolution structures of human Na v 1.7 reveal gating modulation through alpha-pi helical transition of S6 IV.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract:
Nav1.7 represents a preeminent target for next-generation analgesics for its critical role in pain sensation. Here we report a 2.2-Å resolution cryo-EM structure of wild-type (WT) Nav1.7 complexed with the β1 and β2 subunits that reveals several previously indiscernible cytosolic segments. Reprocessing of the cryo-EM data for our reported structures of Nav1.7(E406K) bound to various toxins identifies two distinct conformations of S6IV, one composed of α helical turns only and the other containing a π helical turn in the middle. The structure of ligand-free Nav1.7(E406K), determined at 3.5-Å resolution, is identical to the WT channel, confirming that binding of Huwentoxin IV or Protoxin II to VSDII allosterically induces the α → π transition of S6IV. The local secondary structural shift leads to contraction of the intracellular gate, closure of the fenestration on the interface of repeats I and IV, and rearrangement of the binding site for the fast inactivation motif.
Nav1.7 represents a preeminent target for next-generation analgesics for its critical role in pain sensation. Here we report a 2.2-Å resolution cryo-EM structure of wild-type (WT) Nav1.7 complexed with the β1 and β2 subunits that reveals several previously indiscernible cytosolic segments. Reprocessing of the cryo-EM data for our reported structures of Nav1.7(E406K) bound to various toxins identifies two distinct conformations of S6IV, one composed of α helical turns only and the other containing a π helical turn in the middle. The structure of ligand-free Nav1.7(E406K), determined at 3.5-Å resolution, is identical to the WT channel, confirming that binding of Huwentoxin IV or Protoxin II to VSDII allosterically induces the α → π transition of S6IV. The local secondary structural shift leads to contraction of the intracellular gate, closure of the fenestration on the interface of repeats I and IV, and rearrangement of the binding site for the fast inactivation motif.