{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
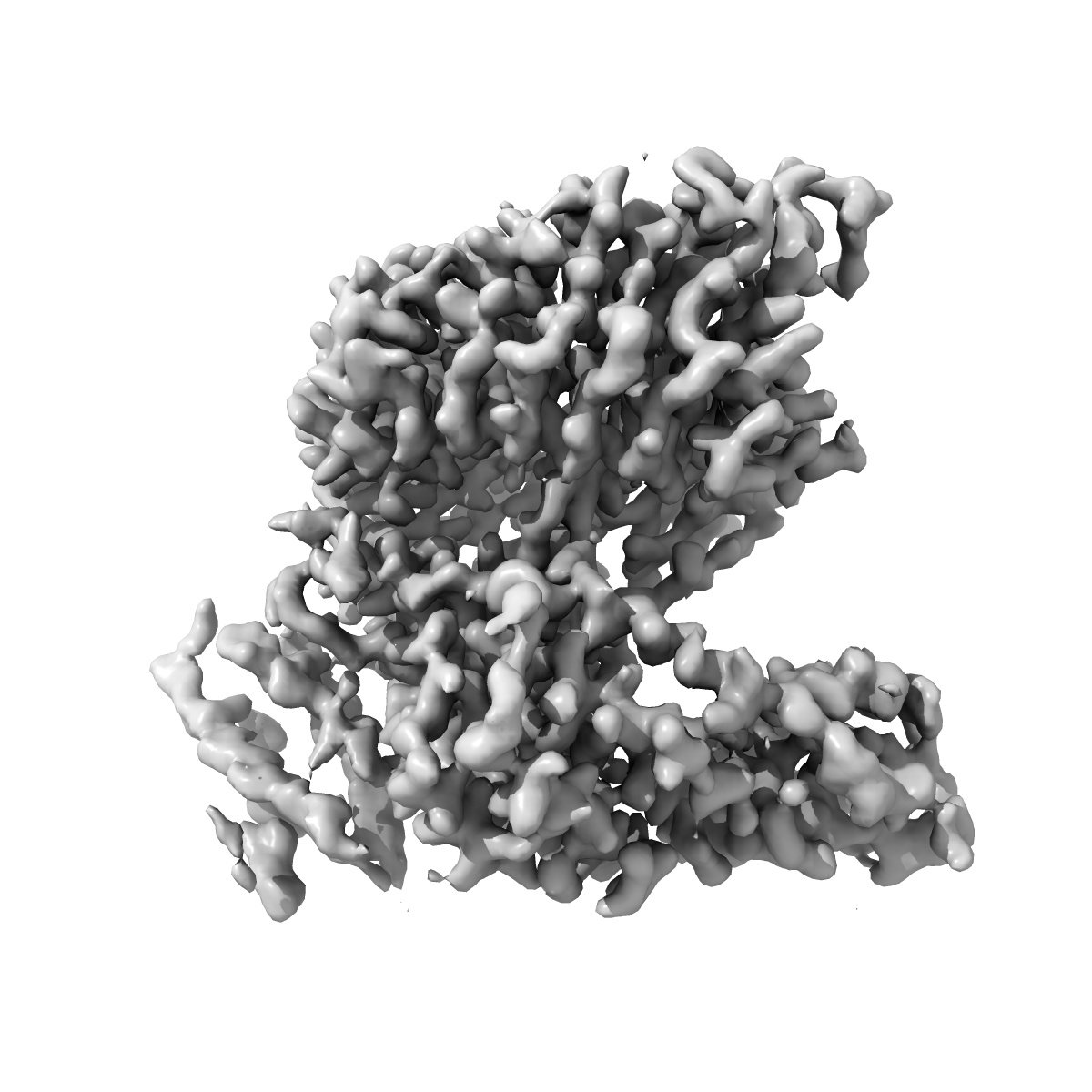
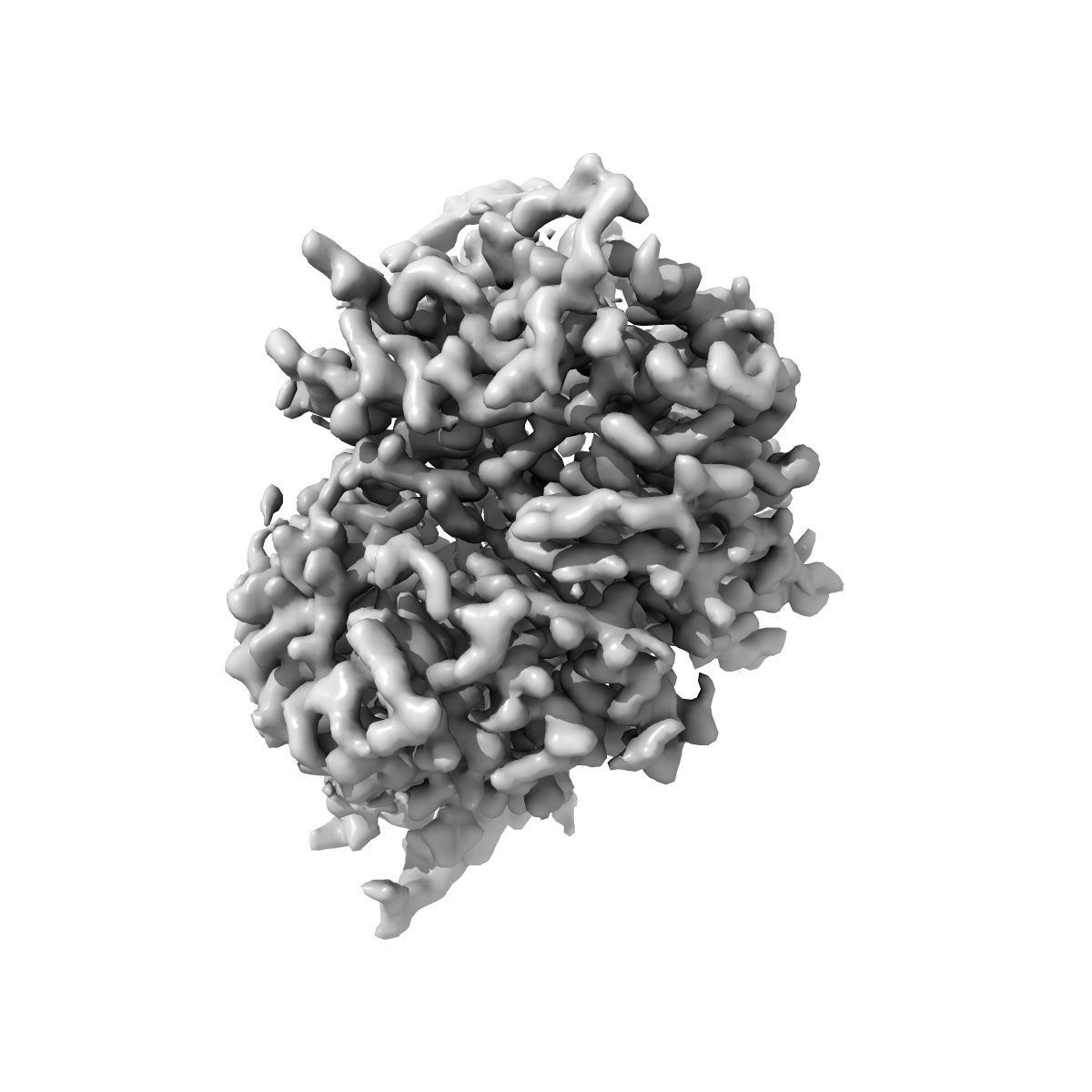
EMD-50863
The barley MLA13-AVRA13 heterodimer
EMD-50863
Single-particle3.8 Å
 Deposition: 03/07/2024
Deposition: 03/07/2024Map released: 12/02/2025
Last modified: 26/02/2025
Sample Organism:
Hordeum vulgare,
Blumeria graminis
Sample: MLA13-AVRA13 heterodimer complex
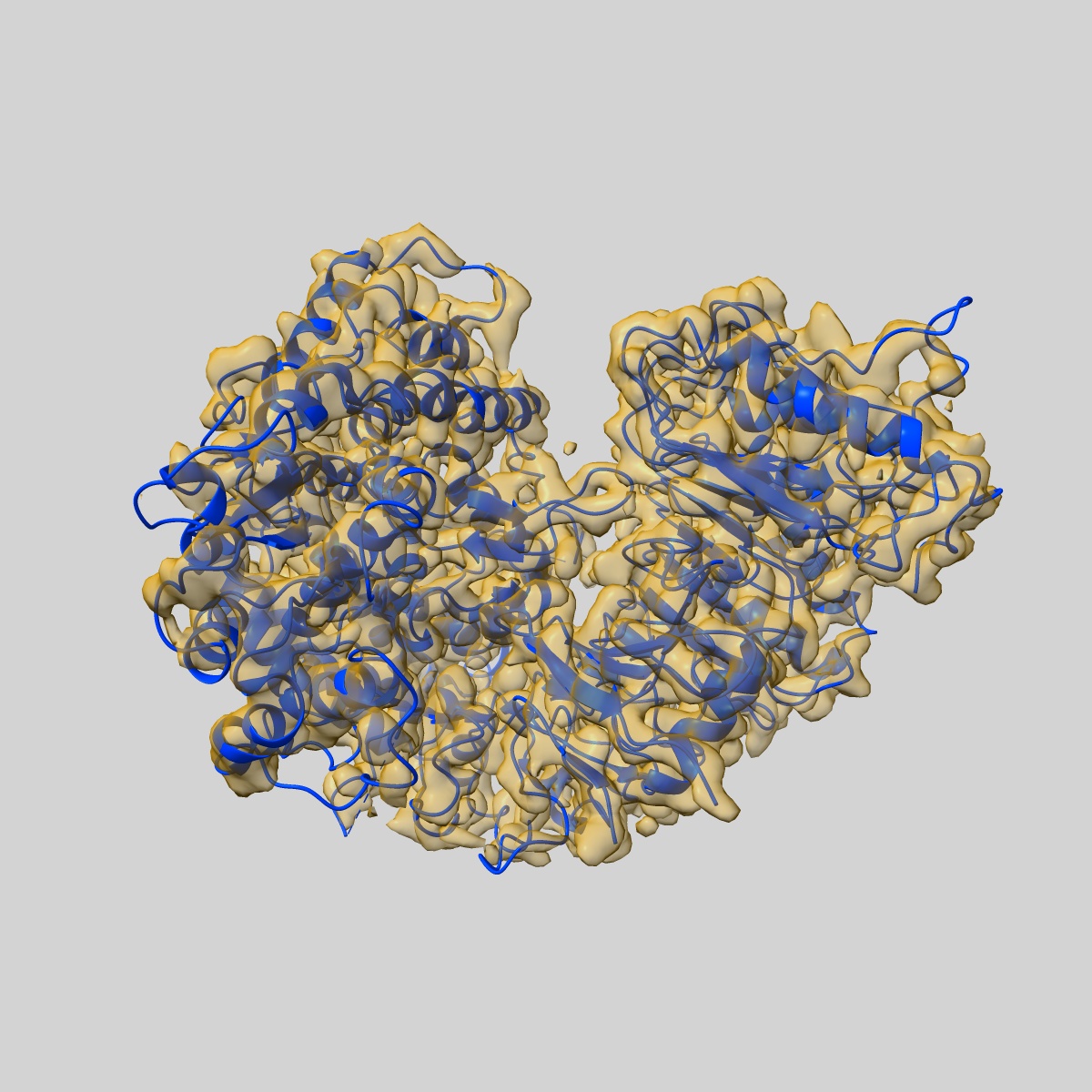
Fitted models: 9fyc (Avg. Q-score: 0.327)
Deposition Authors: Behrmann E ,
Schulze-Lefert P ,
Flores-Ibarra A ,
Lawson AW
,
Schulze-Lefert P ,
Flores-Ibarra A ,
Lawson AW
Sample: MLA13-AVRA13 heterodimer complex
Fitted models: 9fyc (Avg. Q-score: 0.327)
Deposition Authors: Behrmann E
 ,
Schulze-Lefert P ,
Flores-Ibarra A ,
Lawson AW
,
Schulze-Lefert P ,
Flores-Ibarra A ,
Lawson AW
The barley MLA13-AVR A13 heterodimer reveals principles for immunoreceptor recognition of RNase-like powdery mildew effectors.
Lawson AW ,
Flores-Ibarra A ,
Cao Y,
An C,
Neumann U,
Gunkel M,
Saur IML,
Chai J,
Behrmann E ,
Schulze-Lefert P
(2025) EMBO J
,
Flores-Ibarra A ,
Cao Y,
An C,
Neumann U,
Gunkel M,
Saur IML,
Chai J,
Behrmann E ,
Schulze-Lefert P
(2025) EMBO J
Abstract:
Co-evolution between cereals and pathogenic grass powdery mildew fungi is exemplified by sequence diversification of an allelic series of barley resistance genes encoding Mildew Locus A (MLA) nucleotide-binding leucine-rich repeat (NLR) immunoreceptors with an N-terminal coiled-coil domain (CNLs). Each immunoreceptor recognises a matching, strain-specific powdery mildew effector encoded by an avirulence gene (AVRa). We present here the cryo-EM structure of barley MLA13 in complex with its cognate effector AVRA13-1. The effector adopts an RNase-like fold when bound to MLA13 in planta, similar to crystal structures of other RNase-like AVRA effectors unbound to receptors. AVRA13-1 interacts via its basal loops with MLA13 C-terminal leucine-rich repeats (LRRs) and the central winged helix domain (WHD). Co-expression of structure-guided MLA13 and AVRA13-1 substitution variants show that the receptor-effector interface plays an essential role in mediating immunity-associated plant cell death. Furthermore, by combining structural information from the MLA13-AVRA13-1 heterocomplex with sequence alignments of other MLA receptors, we engineered a single amino acid substitution in MLA7 that enables expanded effector detection of AVRA13-1 and the virulent variant AVRA13-V2. In contrast to the pentameric conformation of previously reported effector-activated CNL resistosomes, MLA13 was purified and resolved as a stable heterodimer from an in planta expression system. Our study suggests a common structural principle for RNase-like effector binding to MLAs and highlights the utility of structure-guided engineering of plant immune receptors for broadening their pathogen effector recognition capabilities.
Co-evolution between cereals and pathogenic grass powdery mildew fungi is exemplified by sequence diversification of an allelic series of barley resistance genes encoding Mildew Locus A (MLA) nucleotide-binding leucine-rich repeat (NLR) immunoreceptors with an N-terminal coiled-coil domain (CNLs). Each immunoreceptor recognises a matching, strain-specific powdery mildew effector encoded by an avirulence gene (AVRa). We present here the cryo-EM structure of barley MLA13 in complex with its cognate effector AVRA13-1. The effector adopts an RNase-like fold when bound to MLA13 in planta, similar to crystal structures of other RNase-like AVRA effectors unbound to receptors. AVRA13-1 interacts via its basal loops with MLA13 C-terminal leucine-rich repeats (LRRs) and the central winged helix domain (WHD). Co-expression of structure-guided MLA13 and AVRA13-1 substitution variants show that the receptor-effector interface plays an essential role in mediating immunity-associated plant cell death. Furthermore, by combining structural information from the MLA13-AVRA13-1 heterocomplex with sequence alignments of other MLA receptors, we engineered a single amino acid substitution in MLA7 that enables expanded effector detection of AVRA13-1 and the virulent variant AVRA13-V2. In contrast to the pentameric conformation of previously reported effector-activated CNL resistosomes, MLA13 was purified and resolved as a stable heterodimer from an in planta expression system. Our study suggests a common structural principle for RNase-like effector binding to MLAs and highlights the utility of structure-guided engineering of plant immune receptors for broadening their pathogen effector recognition capabilities.