{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
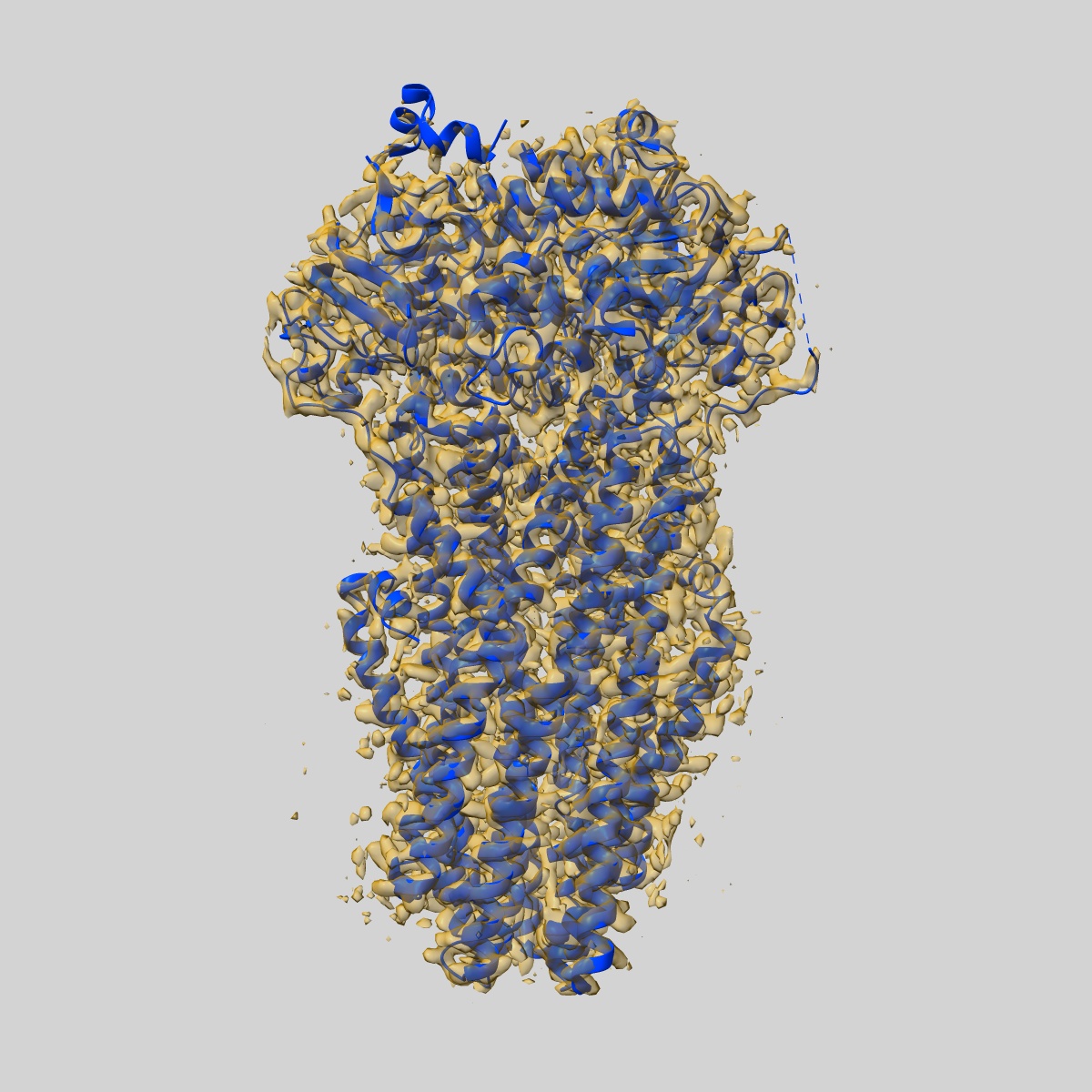
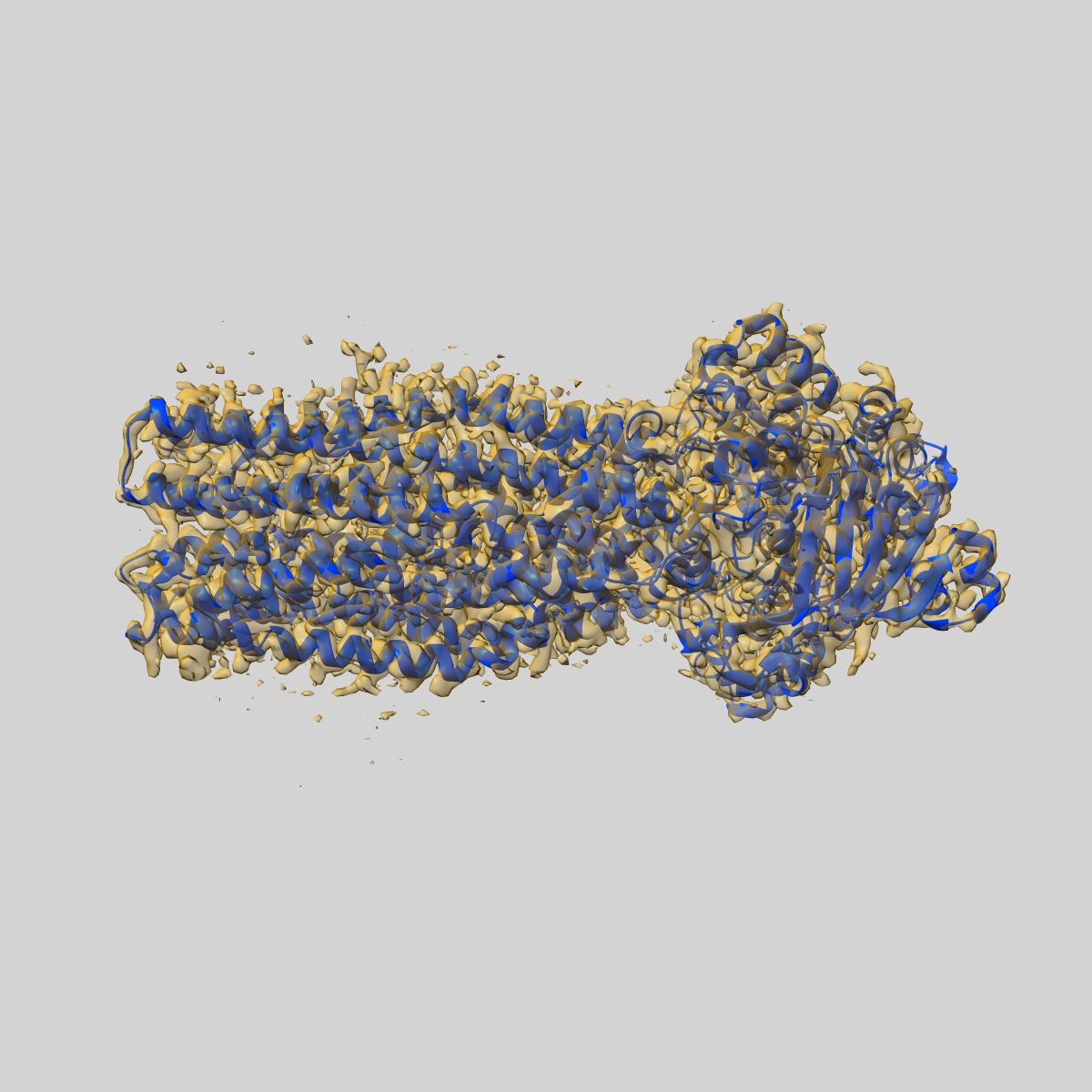
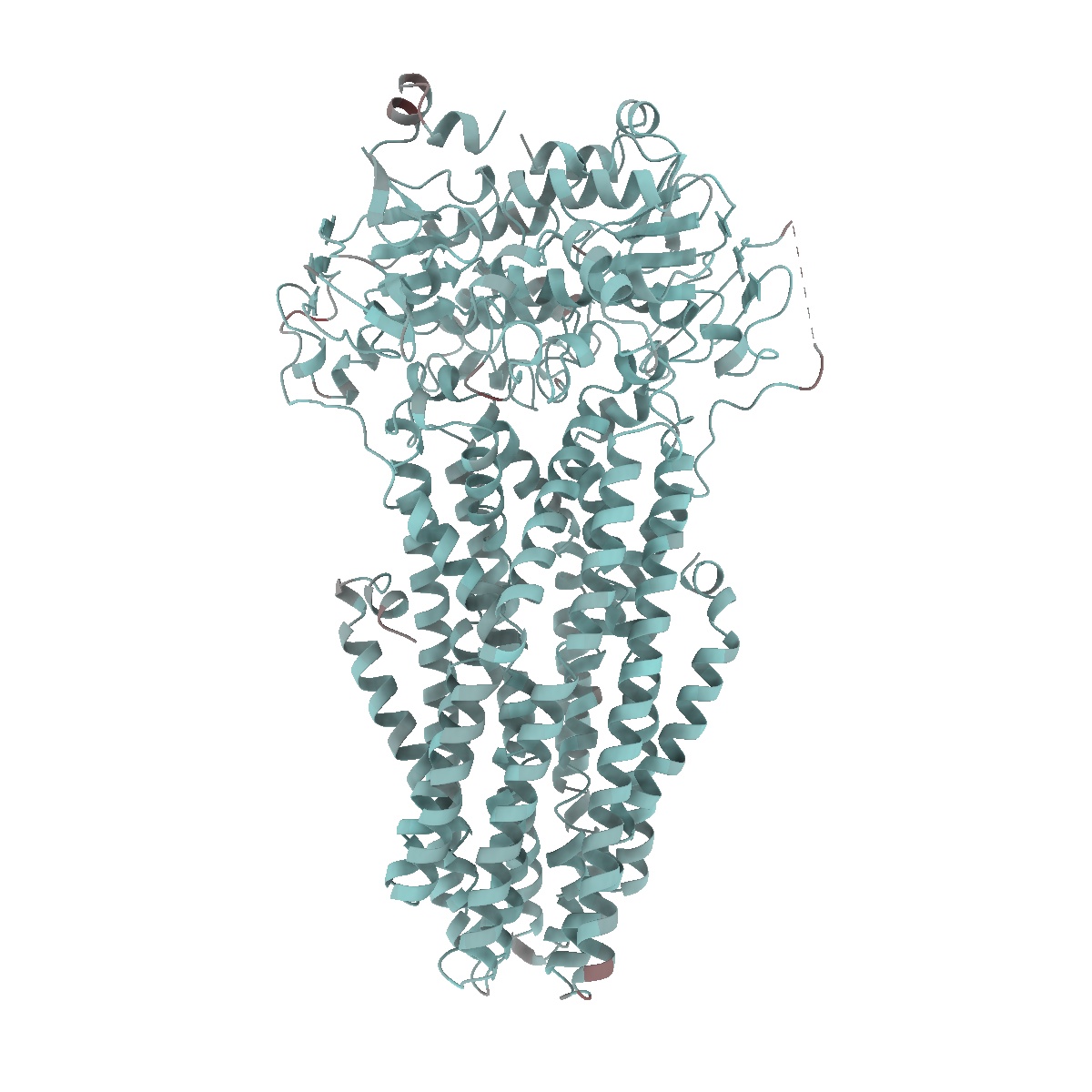
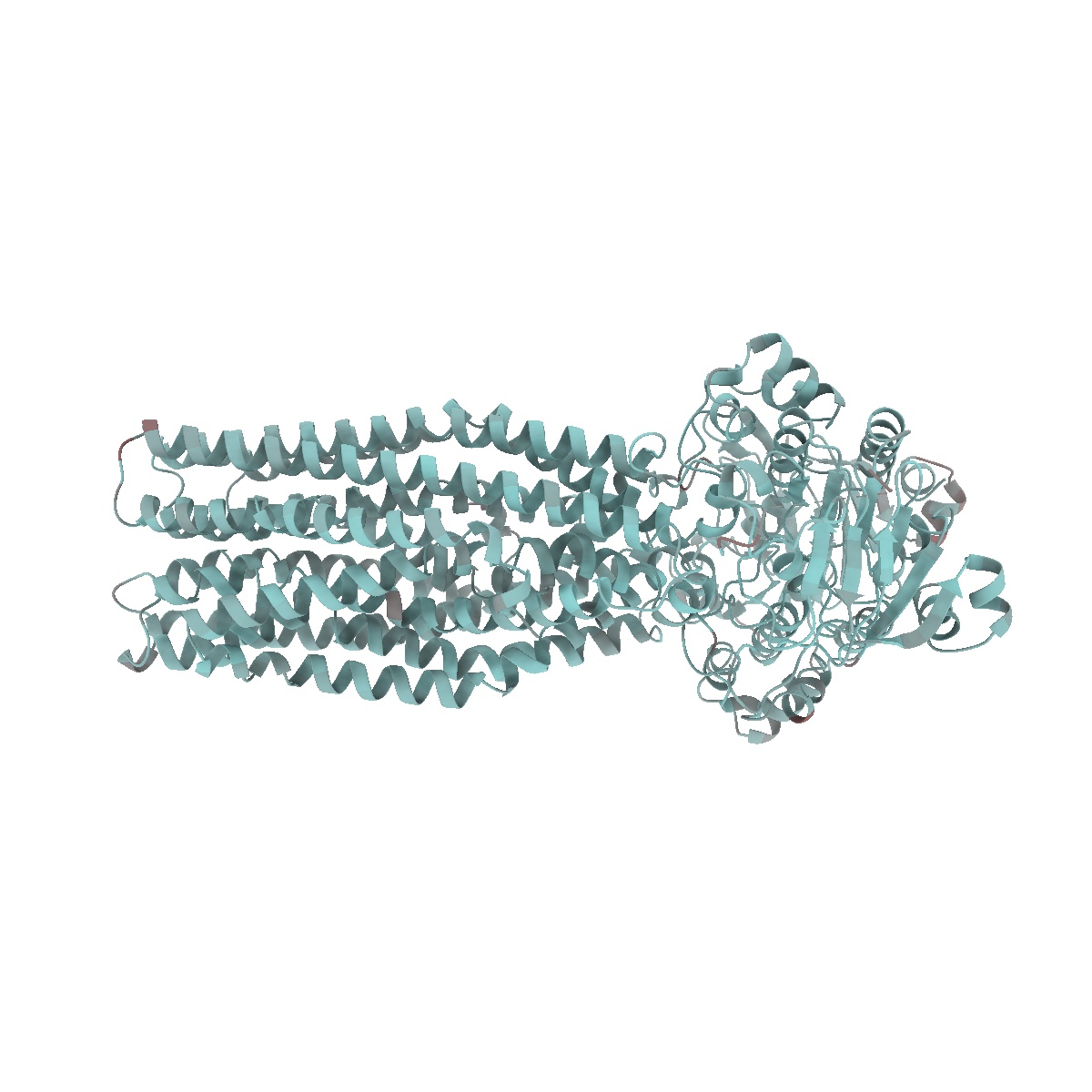
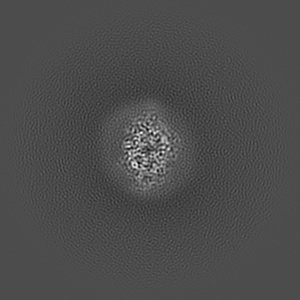
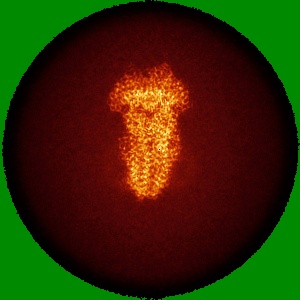
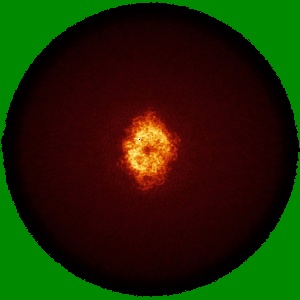
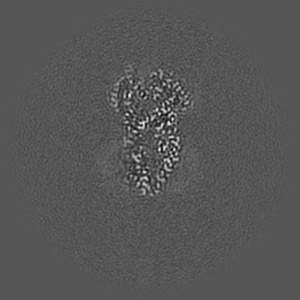
EMD-50980
Cryo-EM structure of IrtAB 2xEQ mutant in outward-occluded state in nanodisc
EMD-50980
Single-particle2.5 Å
 Deposition: 11/07/2024
Deposition: 11/07/2024Map released: 18/12/2024
Last modified: 12/02/2025
Sample Organism:
Mycolicibacterium thermoresistibile ATCC 19527
Sample: IrtAB
Fitted models: 9g2p (Avg. Q-score: 0.64)
Deposition Authors: Gonda I ,
Seeger MA
,
Seeger MA
Sample: IrtAB
Fitted models: 9g2p (Avg. Q-score: 0.64)
Deposition Authors: Gonda I
 ,
Seeger MA
,
Seeger MA
The mycobacterial ABC transporter IrtAB employs a membrane-facing crevice for siderophore-mediated iron uptake.
Gonda I ,
Sorrentino S,
Galazzo L ,
Lichti NP ,
Arnold FM,
Mehdipour AR,
Bordignon E ,
Seeger MA
(2025) Nat Commun , 16 , 1133 - 1133
,
Sorrentino S,
Galazzo L ,
Lichti NP ,
Arnold FM,
Mehdipour AR,
Bordignon E ,
Seeger MA
(2025) Nat Commun , 16 , 1133 - 1133
Abstract:
The mycobacterial ABC transporter IrtAB features an ABC exporter fold, yet it imports iron-charged siderophores called mycobactins. Here, we present extensive cryo-EM analyses and DEER measurements, revealing that IrtAB alternates between an inward-facing and an outward-occluded conformation, but does not sample an outward-facing conformation. When IrtAB is locked in its outward-occluded conformation in nanodiscs, mycobactin is bound in the middle of the lipid bilayer at a membrane-facing crevice opening at the heterodimeric interface. Mutations introduced at the crevice abrogate mycobactin import and in corresponding structures, the crevice is collapsed. A conserved triple histidine motif coordinating a zinc ion is present below the mycobactin binding site. Substitution of these histidine residues with alanine results in a decoupled transporter, which hydrolyzes ATP, but lost its capacity to import mycobactins. Our data suggest that IrtAB imports mycobactin via a credit-card mechanism in a transport cycle that is coupled to the presence of zinc.
The mycobacterial ABC transporter IrtAB features an ABC exporter fold, yet it imports iron-charged siderophores called mycobactins. Here, we present extensive cryo-EM analyses and DEER measurements, revealing that IrtAB alternates between an inward-facing and an outward-occluded conformation, but does not sample an outward-facing conformation. When IrtAB is locked in its outward-occluded conformation in nanodiscs, mycobactin is bound in the middle of the lipid bilayer at a membrane-facing crevice opening at the heterodimeric interface. Mutations introduced at the crevice abrogate mycobactin import and in corresponding structures, the crevice is collapsed. A conserved triple histidine motif coordinating a zinc ion is present below the mycobactin binding site. Substitution of these histidine residues with alanine results in a decoupled transporter, which hydrolyzes ATP, but lost its capacity to import mycobactins. Our data suggest that IrtAB imports mycobactin via a credit-card mechanism in a transport cycle that is coupled to the presence of zinc.