Phosphoserine phosphatase
Phosphoserine phosphatase (PSP) hydrolyses off the phosphoryl group of phospho-L-serine (PLS) and are apart of the haloalkanoic acid dehlogenase superfamily (HADSF). This is a rate-limiting step in mammalian serine biosynthesis and may control mammalian brain development via the NMDA class of glutamate receptors.
Reference Protein and Structure
- Sequence
-
Q58989
 (3.1.3.3)
(3.1.3.3)
 (Sequence Homologues)
(PDB Homologues)
(Sequence Homologues)
(PDB Homologues)
- Biological species
-
Methanocaldococcus jannaschii DSM 2661 (Archaea)
- PDB
-
1l7n
- TRANSITION STATE ANALOGUE OF PHOSPHOSERINE PHOSPHATASE (ALUMINUM FLUORIDE COMPLEX)
(1.8 Å)



- Catalytic CATH Domains
-
3.40.50.1000
 (see all for 1l7n)
(see all for 1l7n)
- Cofactors
- Magnesium(2+) (1)


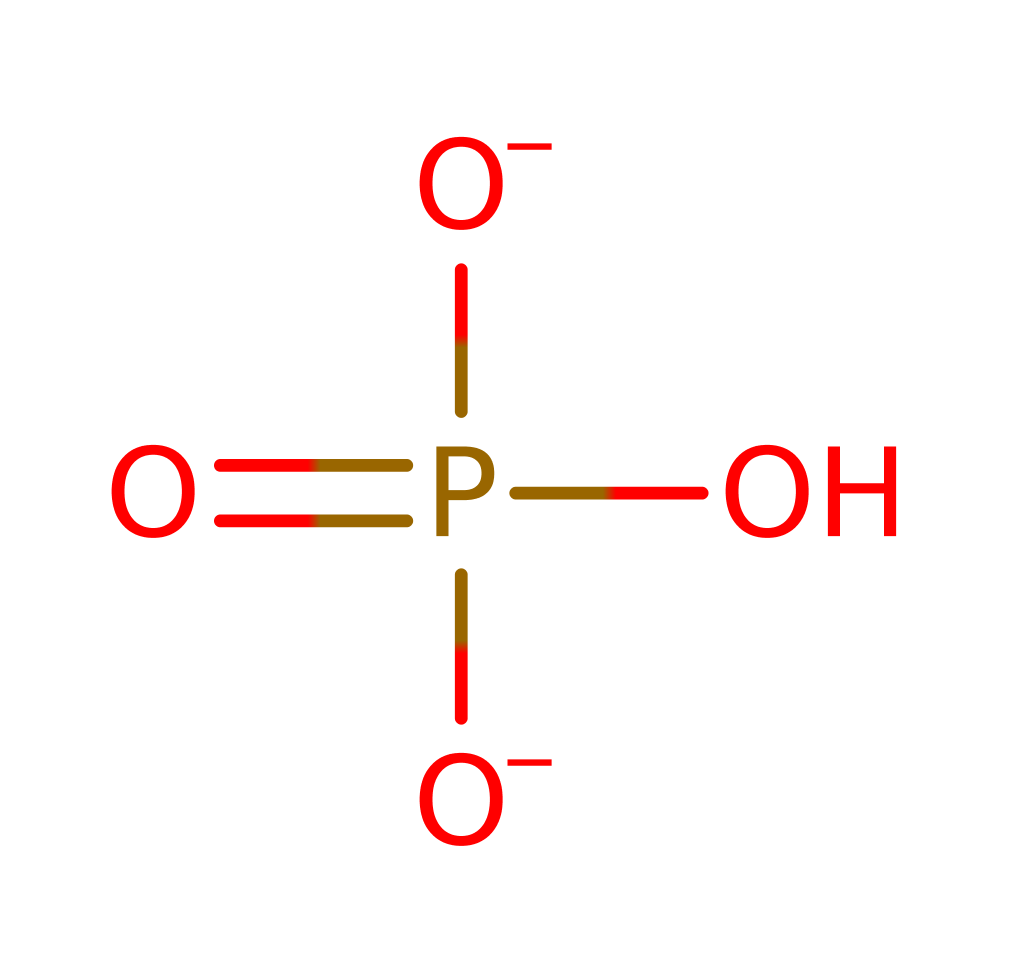

Enzyme Mechanism
Introduction
The basic mechanism is transfer of the phosphoryl group from substrate to Asp 11, and then hydrolysis of the phosphoenzyme intermediate. Whether phosphoryl transfer is primarily associative or dissociative is not yet known and further research is being carried out. The oxyanion hole comprises Mg(II), Lys 144, Asp 171, and the main chain NH groups of Phe 12, Asp 13 and Gly 100. Asp 11 acts as a nucleophile and attacks the phosphorus of the substrate to generate a pentavalent transition state, stabilised by the oxyanion hole. Meanwhile, Asp 13 protonates the serine substrate hydroxyl leaving group resulting in a phosphoenzyme intermediate. A water molecule is activated by Asp 13 acting as a general base and the nucleophile attacks the phosphate group on the intermediate where a second transition state forms, stabilised by the oxyanion hole. Inorganic phosphate is formed and dephosphorylated Asp 11 is regenerated. Aspartate is able to be an ideal nucleophile and leaving group. Additionally, Glu 20 and the cofactor magnesium play an important role in orienting the substrates (magnesium also stabilising the transition states formed during the reaction).
Catalytic Residues Roles
| UniProt | PDB* (1l7n) | ||
| Asp13 | Asp13A | Asp 13 (side chain carboxylic acid / carboxylate) acts as an acid / base catalyst, protonating the serine product and deprotonating water. Asp 13 (main chain NH) is part of the oxyanion hole, activating substrate and phosphoenzyme towards nucleophilic attack and stabilising the charge on the transition states. |
metal ligand, proton acceptor, proton donor |
| Gly100 (main-N) | Gly100A (main-N) | Gly 100 (main chain NH) is part of the oxyanion hole, activating substrate and phosphoenzyme towards nucleophilic attack and stabilising the charge on the transition states. | electrostatic stabiliser |
| Phe12 (main-N) | Phe12A (main-N) | Phe 12 (main chain NH) is part of the oxyanion hole, activating substrate and phosphoenzyme towards nucleophilic attack and stabilising the charge on the transition states. | electrostatic stabiliser |
| Asp11 | Asp11A | Asp 11 is the nucleophile that attacks the substrate phosphorus that is also coordinated to a magnesium ion, phosphorylating itself and releasing the product. | covalently attached, nucleophile, nucleofuge, metal ligand |
| Lys144 | Lys144A | Lys 144 is part of the oxyanion hole, activating substrate and phosphoenzyme towards nucleophilic attack and stabilising the charge on the transition states. | electrostatic stabiliser |
| Asp171 | Asp171A | Asp 171 is part of the oxyanion hole, activating substrate and phosphoenzyme towards nucleophilic attack and stabilising the charge on the transition states. | metal ligand, electrostatic stabiliser |
Chemical Components
proton transfer, bimolecular nucleophilic substitution, overall reactant used, intermediate formation, enzyme-substrate complex formation, dephosphorylation, intermediate terminated, native state of enzyme regenerated, overall product formed, enzyme-substrate complex cleavageReferences
- Wang W et al. (2002), J Mol Biol, 319, 421-431. Structural Characterization of the Reaction Pathway in Phosphoserine Phosphatase: Crystallographic “snapshots” of Intermediate States. DOI:10.1016/s0022-2836(02)00324-8. PMID:12051918.
- Seifried A et al. (2013), FEBS J, 280, 549-571. Human HAD phosphatases: structure, mechanism, and roles in health and disease. DOI:10.1111/j.1742-4658.2012.08633.x. PMID:22607316.
- Allen KN et al. (2009), Curr Opin Struct Biol, 19, 658-665. Markers of fitness in a successful enzyme superfamily. DOI:10.1016/j.sbi.2009.09.008. PMID:19889535.
- Collet JF et al. (1999), J Biol Chem, 274, 33985-33990. Mechanistic Studies of Phosphoserine Phosphatase, an Enzyme Related to P-type ATPases. DOI:10.1074/jbc.274.48.33985. PMID:10567362.

Step 1. Asp 11 acts as a nucleophile and attacks the phosphorus of the substrate to form a phosphoenzyme intermediate. Asp 13 protonates the serine substrate hydroxyl leaving group.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Phe12A (main-N) | electrostatic stabiliser |
| Gly100A (main-N) | electrostatic stabiliser |
| Lys144A | electrostatic stabiliser |
| Asp171A | electrostatic stabiliser |
| Asp11A | metal ligand, covalently attached |
| Asp13A | metal ligand |
| Asp171A | metal ligand |
| Asp11A | nucleophile |
| Asp13A | proton donor |
Chemical Components
proton transfer, ingold: bimolecular nucleophilic substitution, overall reactant used, intermediate formation, enzyme-substrate complex formation, dephosphorylation
Step 2. A water molecule is activated by Asp 13 acting as a general base, releasing the inorganic phosphate from the Asp 11.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Asp11A | metal ligand |
| Phe12A (main-N) | electrostatic stabiliser |
| Gly100A (main-N) | electrostatic stabiliser |
| Lys144A | electrostatic stabiliser |
| Asp171A | electrostatic stabiliser |
| Asp13A | metal ligand |
| Asp171A | metal ligand |
| Asp13A | proton acceptor |
| Asp11A | nucleofuge |
 Download:
Download: