{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
EMD-18581
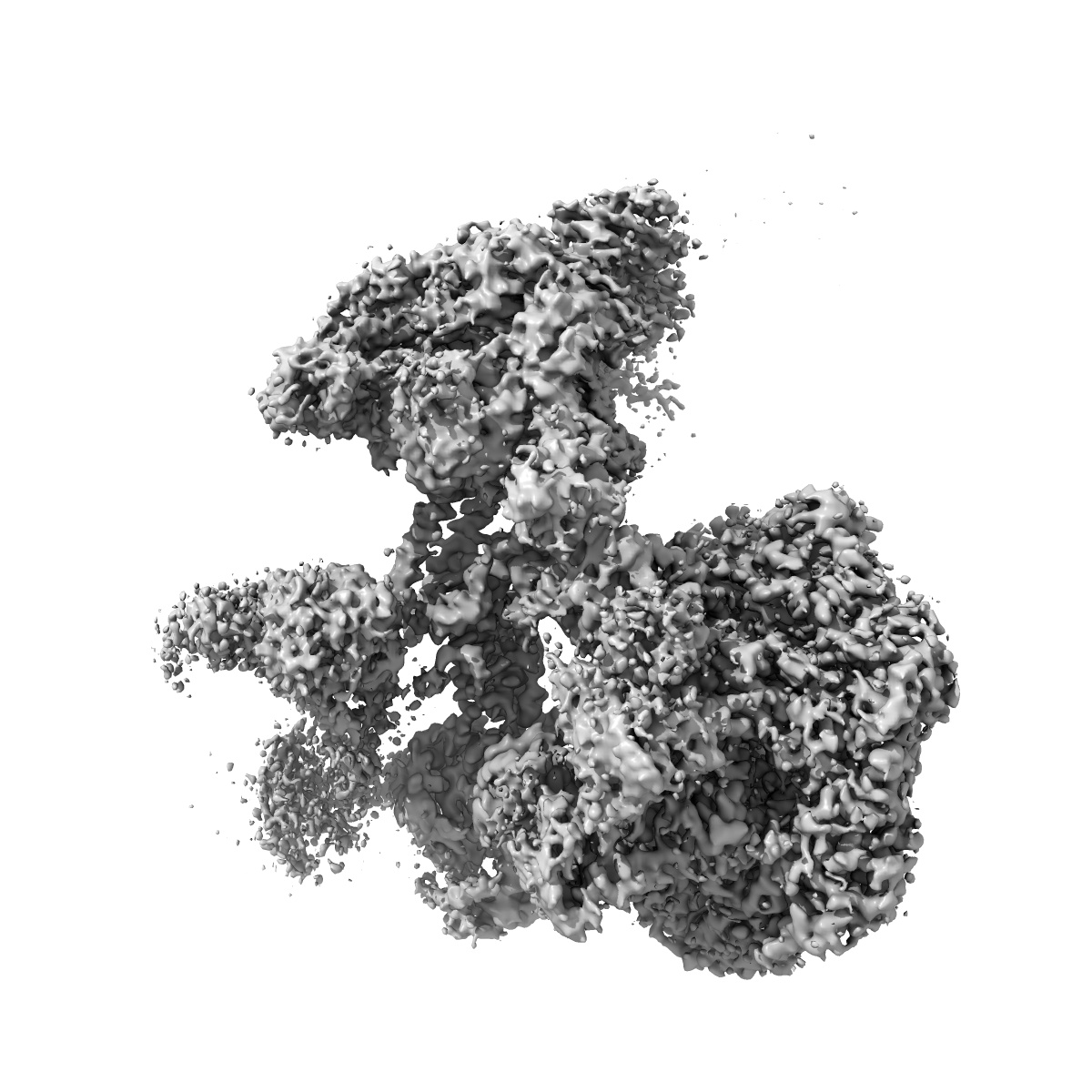
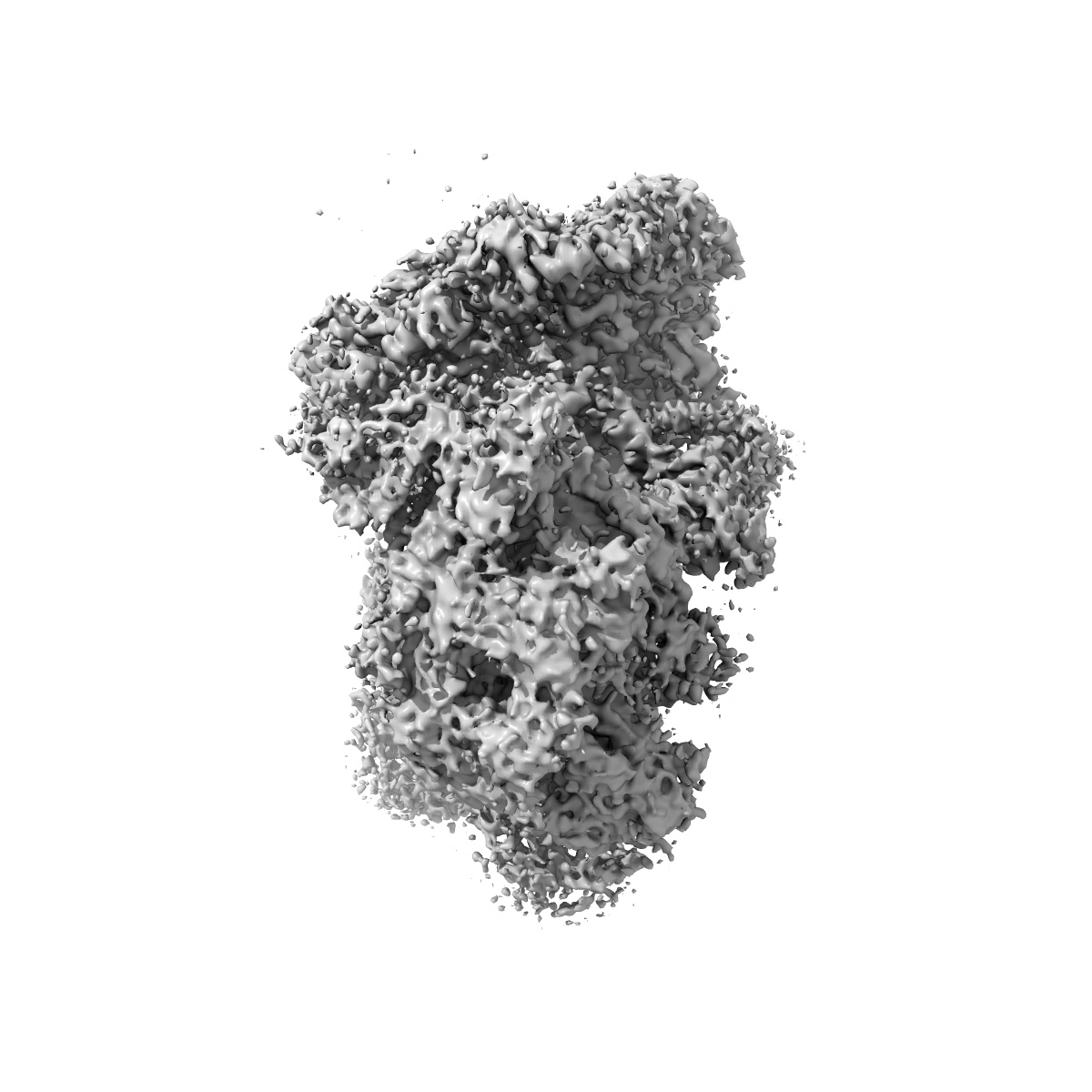
Human Tip60 complex
EMD-18581
Single-particle2.52 Å
 Deposition: 03/10/2023
Deposition: 03/10/2023Map released: 07/08/2024
Last modified: 04/12/2024
Sample Organism:
Homo sapiens
Sample: Tip60 complex
Deposition Authors: Li C, Smirnova E, Schnitzler C, Crucifix C, Concordet JP, Brion A, Poterszman A ,
Schultz P ,
Papai G ,
Ben-Shem A
,
Schultz P ,
Papai G ,
Ben-Shem A
Sample: Tip60 complex
Deposition Authors: Li C, Smirnova E, Schnitzler C, Crucifix C, Concordet JP, Brion A, Poterszman A
 ,
Schultz P ,
Papai G ,
Ben-Shem A
,
Schultz P ,
Papai G ,
Ben-Shem A
Structure of the human TIP60-C histone exchange and acetyltransferase complex.
Li C,
Smirnova E,
Schnitzler C,
Crucifix C,
Concordet JP,
Brion A,
Poterszman A ,
Schultz P ,
Papai G ,
Ben-Shem A
(2024) Nature , 635 , 764 - 769
,
Schultz P ,
Papai G ,
Ben-Shem A
(2024) Nature , 635 , 764 - 769
Abstract:
Chromatin structure is a key regulator of DNA transcription, replication and repair1. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two enzymatic activities: ATP-dependent exchange of histone H2A-H2B for H2A.Z-H2B, and histone acetylation. In yeast, however, these activities are performed by two independent complexes-SWR1 and NuA4, respectively2,3. How the activities of the two complexes are merged into one supercomplex in humans, and what this association entails for the structure and mechanism of the proteins and their recruitment to chromatin, are unknown. Here we describe the structure of the endogenous human TIP60-C. We find a three-lobed architecture composed of SWR1-like (SWR1L) and NuA4-like (NuA4L) parts, which associate with a TRRAP activator-binding module. The huge EP400 subunit contains the ATPase motor, traverses the junction between SWR1L and NuA4L twice and constitutes the scaffold of the three-lobed architecture. NuA4L is completely rearranged compared with its yeast counterpart. TRRAP is flexibly tethered to NuA4L-in stark contrast to its robust connection to the completely opposite side of NuA4 in yeast4-7. A modelled nucleosome bound to SWR1L, supported by tests of TIP60-C activity, suggests that some aspects of the histone exchange mechanism diverge from what is seen in yeast8,9. Furthermore, a fixed actin module (as opposed to the mobile actin subcomplex in SWR1; ref. 8), the flexibility of TRRAP and the weak effect of extranucleosomal DNA on exchange activity lead to a different, activator-based mode of enlisting TIP60-C to chromatin.
Chromatin structure is a key regulator of DNA transcription, replication and repair1. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two enzymatic activities: ATP-dependent exchange of histone H2A-H2B for H2A.Z-H2B, and histone acetylation. In yeast, however, these activities are performed by two independent complexes-SWR1 and NuA4, respectively2,3. How the activities of the two complexes are merged into one supercomplex in humans, and what this association entails for the structure and mechanism of the proteins and their recruitment to chromatin, are unknown. Here we describe the structure of the endogenous human TIP60-C. We find a three-lobed architecture composed of SWR1-like (SWR1L) and NuA4-like (NuA4L) parts, which associate with a TRRAP activator-binding module. The huge EP400 subunit contains the ATPase motor, traverses the junction between SWR1L and NuA4L twice and constitutes the scaffold of the three-lobed architecture. NuA4L is completely rearranged compared with its yeast counterpart. TRRAP is flexibly tethered to NuA4L-in stark contrast to its robust connection to the completely opposite side of NuA4 in yeast4-7. A modelled nucleosome bound to SWR1L, supported by tests of TIP60-C activity, suggests that some aspects of the histone exchange mechanism diverge from what is seen in yeast8,9. Furthermore, a fixed actin module (as opposed to the mobile actin subcomplex in SWR1; ref. 8), the flexibility of TRRAP and the weak effect of extranucleosomal DNA on exchange activity lead to a different, activator-based mode of enlisting TIP60-C to chromatin.