
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
EMD-25529
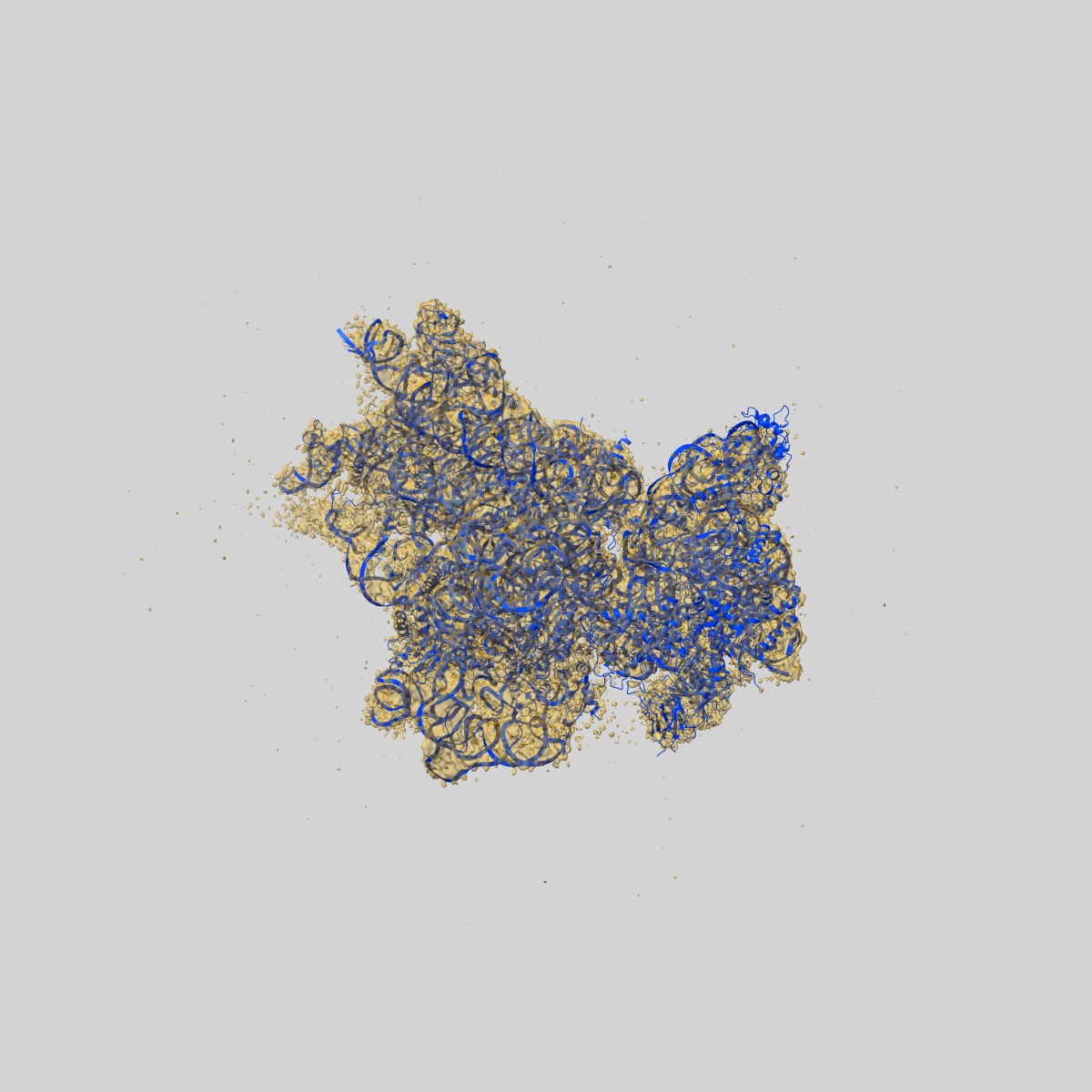
Structure of the HCV IRES binding to the 40S ribosomal subunit, closed conformation. Structure 3(delta dII)
EMD-25529
Single-particle4.5 Å
 Deposition: 25/11/2021
Deposition: 25/11/2021Map released: 13/07/2022
Last modified: 05/06/2024
Sample Organism:
Oryctolagus cuniculus,
Hepacivirus C
Sample: 40S ribosomal small subunit with HCV IRES binding
Fitted models: 7syi (Avg. Q-score: 0.323)
Deposition Authors: Brown ZP ,
Abaeva IS ,
De S ,
Hellen CUT ,
Pestova TV ,
Frank J
,
Abaeva IS ,
De S ,
Hellen CUT ,
Pestova TV ,
Frank J
Sample: 40S ribosomal small subunit with HCV IRES binding
Fitted models: 7syi (Avg. Q-score: 0.323)
Deposition Authors: Brown ZP
 ,
Abaeva IS ,
De S ,
Hellen CUT ,
Pestova TV ,
Frank J
,
Abaeva IS ,
De S ,
Hellen CUT ,
Pestova TV ,
Frank J
Molecular architecture of 40S translation initiation complexes on the hepatitis C virus IRES.
Brown ZP ,
Abaeva IS ,
De S ,
Hellen CUT ,
Pestova TV ,
Frank J
(2022) EMBO J , 41 , e110581 - e110581
,
Abaeva IS ,
De S ,
Hellen CUT ,
Pestova TV ,
Frank J
(2022) EMBO J , 41 , e110581 - e110581
Abstract:
Hepatitis C virus mRNA contains an internal ribosome entry site (IRES) that mediates end-independent translation initiation, requiring a subset of eukaryotic initiation factors (eIFs). Biochemical studies revealed that direct binding of the IRES to the 40S ribosomal subunit places the initiation codon into the P site, where it base pairs with eIF2-bound Met-tRNAiMet forming a 48S initiation complex. Subsequently, eIF5 and eIF5B mediate subunit joining, yielding an elongation-competent 80S ribosome. Initiation can also proceed without eIF2, in which case Met-tRNAiMet is recruited directly by eIF5B. However, the structures of initiation complexes assembled on the HCV IRES, the transitions between different states, and the accompanying conformational changes have remained unknown. To fill these gaps, we now obtained cryo-EM structures of IRES initiation complexes, at resolutions up to 3.5 Å, that cover all major stages from the initial ribosomal association, through eIF2-containing 48S initiation complexes, to eIF5B-containing complexes immediately prior to subunit joining. These structures provide insights into the dynamic network of 40S/IRES contacts, highlight the role of IRES domain II, and reveal conformational changes that occur during the transition from eIF2- to eIF5B-containing 48S complexes and prepare them for subunit joining.
Hepatitis C virus mRNA contains an internal ribosome entry site (IRES) that mediates end-independent translation initiation, requiring a subset of eukaryotic initiation factors (eIFs). Biochemical studies revealed that direct binding of the IRES to the 40S ribosomal subunit places the initiation codon into the P site, where it base pairs with eIF2-bound Met-tRNAiMet forming a 48S initiation complex. Subsequently, eIF5 and eIF5B mediate subunit joining, yielding an elongation-competent 80S ribosome. Initiation can also proceed without eIF2, in which case Met-tRNAiMet is recruited directly by eIF5B. However, the structures of initiation complexes assembled on the HCV IRES, the transitions between different states, and the accompanying conformational changes have remained unknown. To fill these gaps, we now obtained cryo-EM structures of IRES initiation complexes, at resolutions up to 3.5 Å, that cover all major stages from the initial ribosomal association, through eIF2-containing 48S initiation complexes, to eIF5B-containing complexes immediately prior to subunit joining. These structures provide insights into the dynamic network of 40S/IRES contacts, highlight the role of IRES domain II, and reveal conformational changes that occur during the transition from eIF2- to eIF5B-containing 48S complexes and prepare them for subunit joining.