represent EMBL in Europe.
Tissue biology and disease modelling
Structural biology
Main laboratory
EMBL-EBI: European Bioinformatics Institute
Epigenetics and neurobiology
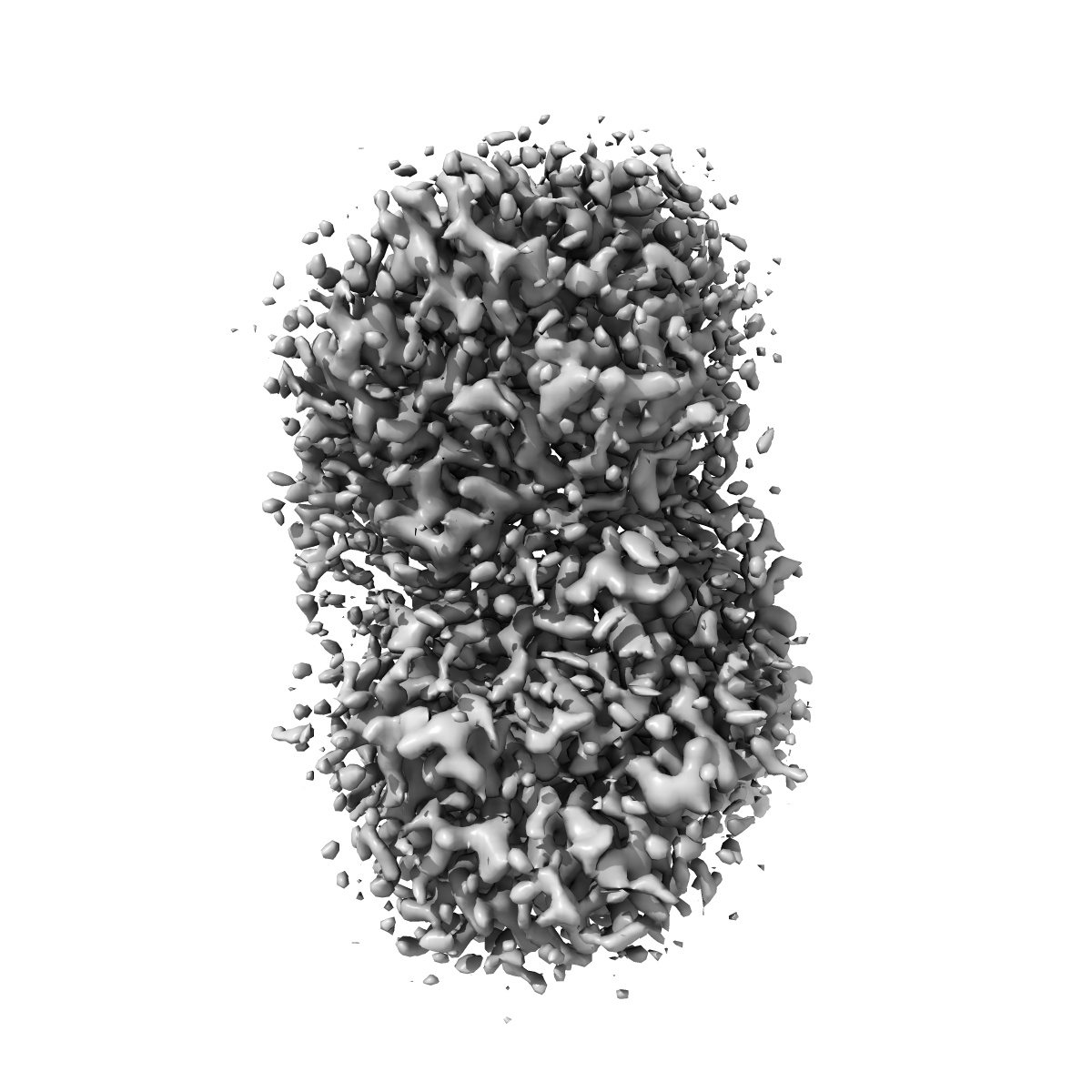
Examples: 1001, Apoferritin, Tomography, Rossmann MG, 5A1A
advanced search
Searching in EMDBHelp
Share EMDB
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 Deposition: 12/01/2024
Deposition: 12/01/2024 ,
Zhao B
,
Zhao B
 ,
Zhao B
,
Zhao B